Romania - Numărul de Profesori la Numărul de Elevi
Securistoidul Pantazi un analfabet funcțional în domeniul economiei declara ritos că "distributivismul este baza marxismului". Mai dobitocule doctrina distributivista a fost creata de un Papă si dezvoltata de un alt Papă
Nimeni nu e mai presus de Lege. Nici măcar Curtean Constituțională
Motivarea judecatorului
Prăbușirea Apple demonstrează că a te gudura pe lângă Trump nu este o garanție a siguranței
Prăbușirea demografică a României se datorează în integralitatea partidelor sistemului care au condus România 35 de ani
Prăbușirea criptomonedelor TerraUS și LUNA s-a datorat unui grup restrâns de speculanți care au pariat "short"
Explicația pe scurt a Tarifelir vamale impuse de Trump și cât de mult ajută economia americană
Rezultatele sondajului Verifield
Donald Trump cere eliberarea lui Marine Le Pen și să fie lăsată să candideze. Nici un cuvânt despre anularea alegerilor și nici despre Călin Georgescu spre satisfacția sorosistilor și a slugilor lor care au dat Lovitura de Stat in România
SpaceX tocmai a făcut un pas mare spre reutilizarea amplificatorului Super Heavy al Starship
ActiveNewx - Bolojan și-a adus la Palat "societatea civilă" SOROS după modelul Iohannis pentru a discuta combaterea ”știrilor false” înainte de alegeri, adică perfecționarea cenzurii
Trăim în URSS. Procurorii CSM si seful PICCJ Alex Florenta cer cercetarea judecătoarei Boboc de la CA Constanta pentru hotararea prin care a stramutat dosarul lui Dan Diaconescu si a desfiintat condamnarea
Elon Musk a criticat vehement verdictul instanței franceze care a blocat candidatura lui Marine Le Pen la alegerile prezidențiale din 2027. Cazul Călin Georgescu a fost din nou Reamintit.
Ca și în cazul lui Călin Georgescu, Regimul Soroșist Macron o interzice pe Marine Le Pen. Le Pen găsită vinovată. Le Pen interzisă să candideze.
La fel ca și Călin Georgescu care avea peste 40% în turul întâi, Marine le Pen conduce detașat in sondaje pentru cursa prezidențială. E momentul ca regimul Neobolsevic Soroșist să o scoată din joc cum a făcut și la noi.
Viktor Orban: "Je suis Marine!". Gunoiul de Viktor Orban de interzicerea lui Călin Georgescu nu a spus nimic desi acesta nu era condamnat deloc. Ce înseamnă să fii lipsit de caracter
Un nou sondaj: Simion pe primul loc, Ponta pe doi. Sondaj Verifield comandat de Nicușor Dan.
Cazul Le Pen, ineligibilitate și executare provizorie: ce spune cu adevărat legea
Sorosistul Securistoid Bănescu noua Față Hidoase a Cenzurii Regimului UEist de la București
Ciutacu alias Saul Bruckner 2 in Scânteia 2 sau Cum sa înjuri in stil bolșevic românii doar pentru că votează cu cine vor ei? Tonomatul puterii pe nume Ciutacu care pupă poala Pucistului Ciolacu și a lui Gold Ponta înjură oribil votanții lui Georgescu
De ce sa înjuri votanții că votează cu cine vor ei? Tonomatul puterii pe nume Ciutacu care pupă la greu poala Pucistului Ciolacul și a lui Gold Corporation Ponta ii înjură oribil pe votanții lui Călin Georgescu.
De ce sa înjuri votanții că votează cu cine vor ei? Tonomatul puterii pe nume Ciutacu care pupă la greu poala Pucistului Ciolacul și a lui Gold Corporation Po ta ii înjură într-un mod oribil pe votanții lui Călin Georgescu.
De ce sa înjuri votanții că votează cu cine vor ei? Tonomatul puterii pe nume Ciutacu care pupă la greu poala Pucistului Ciolacul și a lui Gold Corporation Ponta ii înjură oribil pe votanții lui Georgescu
Ion Cristoiu: Votul din decembrie a fost moral Lumea s-a săturat de "politruceală" Da, inevitabil, ne despărțim de America (referindu-se la puciști)
Reacția Cristelei Georgescu după ce Puciștii care au dat Lovitura de Stat, l-au interzis pe soțul ei: „Nimic nu este pierdut atât timp cât credinţa este în picioare, cât sufletul nu abdică şi cât capul se ridică din nou.”
Adrian Onciu: Cine sunt, de fapt, extremiștii
CURS-ul lui Buble îl urca pe Crin Antonescu pe locul doi. Sondaj CURS pentru alegerile prezidențiale. 59% dintre români cred că direcția țării impregnată de Puciști este greșită
Ca în Botswana, fiecare Institut de sondare are un candidat preferat. SOCIOPOL-ul lui Palada îl urcă pe Ponta pe locul doi.
În sondajul Atlas Intel Soroșiștul Nicușor Dan, e pe locul doi
Pieleanu "face" sondaje "la refefon"
Ca și bolșevicii în 1946, Soroșiștii nu au stare până nu bagă Cenzura peste tot. Lasconi cere închiderea TikTok înaintea prezidenţialelor: "Fake news-ul să fie dat jos". Soroșiștii de la Funky Citizens decid ei de capul lor, ce e "fake news"
Ramurile Cominterniste directe sau prin alianță se susțin între ele. Celebrul Bombonel, alias Adrian Năstase, care a dat petrolul țării și multe alte bunuri naționale pe Nimic, îl susține pe Victor Ponta. Nu ai nenorocit destul țara asta Bombonele?
Subordonat rețelei soroșiste ca și Nicușor Dan, Bilderbergul Merz pune biciul pe germani. Germania se înarmează până în dinți și vrea să taie pensii, ajutoarele sociale și subvenția pentru energie
Viktor Orbán: "Nu mai puteam să stăm cu mâinile în sân şi să privim cum lanţurile de magazine jefuiesc oamenii". Vă amintiți cum au sărit Soroșiștii pe Călin Georgescu când ne-a spus că Multinaționalele ne jefuiesc? De ce AUR și Simion TAC pe această temă?
Calin Georgescu a avut dreptate când a arătat jaful multinaționalelor. Bulgaria înființează lanțuri de magazine cu adaos 10%, pentru a împiedeca multinaționalele să jefuiască populația. Puciștii de la noi sunt pe-o mână cu multinaționalele
Editorial Cornel Dinu. SMINTEALA clasei politice înaintea unor alegeri prezidențiale compromise din start
În 35 de ani de Soroșism, Familia Tradițională a fost distrusă. UE A AJUNS LA 700.000 DE DIVORȚURI Rata divorțurilor s-a triplat. Unu din doi copii se nasc in afara căsătoriei. Înțelegeți de ce Călin Georgescu a spus că Familia trebuie protejata?
Tăcerea democrației: strigătul unei națiuni pentru libertate<
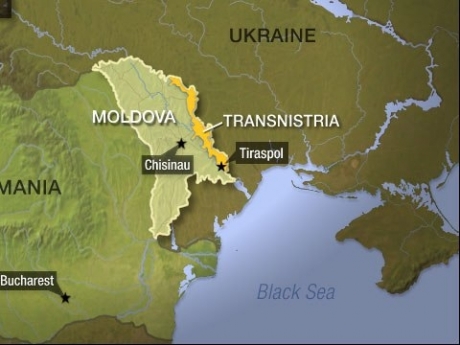
Așadar dacă pacea se va încheia Nikolaev și Odessa vor rămâne în Ucraina, în sfera de influență Americană. Putin nu va revendica Odesa dacă americanii recunosc Crimeea, Donețk, Lugansk, Zaporojie și Herson ca părți ale Rusiei
Luptătorul din Arenă
Vă sună cunoscut? Am văzut în România. Primarul Istanbulului, progresistul Imamoglu, arestat Principalul adversar al lui Erdogan, Imamoglu își anunțase candidatura la președinție in 2028
Mișcarea Suveranistă e împărțită între "Turul Doi Inapoi" și "Sa mergem înainte cu Simion". Simion și Ana Maria Gavrilă trebuie sa sustina Rolul Important a lui Călin Georgescu în Statul Român dacă vor să castige
Micul Stalinist Nicușor Soros Dan ne spune că a&i lua unui om dreptul de a candida și milioanelor de români dreptul de a-l vota fără o condamnare "e o decizie corectă". Poate in URSS-ul lui Lenin, plicușor cu bani de la Soros
Așadar după mintea actualului Stat Soroșist și a cohortelor de sorosisti care aplaudă: Democrația înseamnă să anulezi alegerile, să-l interzicem, să-i facem dosar penal și să-l băgăm la zdup pe cel CARE A CÂȘTIGAT ALEGERILE PRIN VOT
România propusă de Călin Georgescu vs România puciștilor dirijați de progresista UE. Iată cum arată România Puciștilor : Șomajul în rândul tinerilor e cel mai mare din UE. La țară a ajuns peste 30%
Bilderbergul Macron: UE poate sprijini Ucraina fără aprobarea Rusiei trimițând mii de soldați din fiecare țară a UE.
Libertatea de exprimare a murit. Lasconi de la USR cere și mai multă CENZURĂ. Soroșismului Neobolsevic devine tot mai Totalitar. "cel puțin 2 televiziuni își fac de cap" "avem proiect de Lege" urlă Lasconi
Adrian Onciu - Active News : Continuă telenovela nord-coreeană de admitere sau respingere a candidaților la prezidențiale în funcție de cum dictează autorii loviturii de stat.










:format(webp):quality(80)/https://romaniatv.net/wp-content/uploads/2014/08/520364_dinu_publimedia_cosmin_motei_56904200.jpg)
































































Comentarii:
Adauga Comentariu